

Reservoir formation mechanism of the microbialite in the Fengjiawan Formation of the Mesoproterozoic Jixian System, southern Ordos Basin
-
摘要:
在鄂尔多斯盆地南缘中元古界蓟县系冯家湾组硅质叠层石和鲕粒岩中,存在白云石及其溶蚀形成的孔洞。分布特征与发育规模明显受富有机质组构控制的白云石和溶蚀孔洞是基质孔隙发育的关键。显微薄片、扫描电镜、阴极发光、大面积拼接成像及微量元素原位成像等分析显示,大量自形白云石晶体分布于叠层石的暗色纹层与鲕粒的暗色圈层中,且晶体周缘普见缝隙的存在。部分白云石发生了溶蚀,形成晶体铸模孔并成为暗色组构中孔洞的基本构成单元。白云石的形成与随后的溶蚀均发生在富有机质组构内部。组构内受微生物因素影响的成岩物质的富集,以及随之产生的环境酸碱度变化,共同促进了白云石的发育与溶解过程。上述现象和认识可以为前寒武系部分白云岩的成因和可能与微生物因素有关的成岩、成储研究提供一些参考借鉴。
Abstract:Dolomite crystals and related pores and vugs, evidently controlled by organic-rich fabrics, can be found in the siliceous stromatolites and oolites in the Fengjiawan Formation of the Mesoproterozoic Jixian System, located in the southern Ordos Basin. With the application of microscope, scanning electron microscope (SEM), cathodoluminescence, large-area SEM image acquisition, and in-situ trace element imaging, it can be observed that many euhedral dolomite crystals are concentrated within the dark laminae of the stromatolites and the dark spheres of the ooids, with gaps present on the crystal peripheries. A considerable number of these dolomite crystals have undergone dissolution, leading to the formation of moldic pores and vugs, which are the dominant matrix pores in the stromatolites and oolites in the study area. Dolomite formation and dissolution were confined to the organic-rich fabrics. The microenvironment within these fabrics was influenced by microbial factors, with the enrichment of necessary diagenetic substances and changes in pH. These phenomena and scientific understandings may have some referential value for the genesis of some Precambrian dolomites and microbial-related diagenesis, which are of great significance for petroleum geology.
-
Keywords:
- Ordos Basin /
- Fengjiawan Formation /
- microbialite /
- dolomite /
- dissolution
-
0. 引言
中国前寒武系海相碳酸盐岩广泛分布于四川盆地、塔里木盆地、华北地台等地区,虽然时代古老且很多埋藏较深,但仍具有可观的油气储集潜力,在能源紧缺的严峻形势下,成为潜在的油气大发现的勘探新领域(胡安平等,2021)。前寒武纪生态系以包括细菌、疑源类、真核藻类等在内的微生物占统治地位为特征,导致微生物碳酸盐岩(简称微生物岩)在太古宙和元古宙尤其是中元古代大规模发育,形成以叠层石为代表的多种类型的生物−沉积构造,构成前寒武系碳酸盐岩的主要组成部分(Riding,1991,2000,2006;Johnston et al.,2009;Bosence et al.,2015;Peters et al.,2017)。鄂尔多斯盆地中元古界蓟县系发育有大量微生物岩。近年来,随着华北地块陆续发现中元古界油气显示,以及甘肃崇信地区蓟县系古油藏沥青的发现(马赛,2020),蓟县系成为资源潜力逐渐显现的潜在勘探层系。
前人针对微生物的生命活动在前寒武系碳酸盐岩形成过程中所发挥的造岩作用开展了大量研究。例如,叠层石是以蓝细菌为主的微生物周期性的生长(包含机械性粘结捕获和微生物诱导矿化)与碳酸钙沉淀物交替发育、共同构建的产物(Riding,1991,2008;Reid et al.,2000;Pace et al.,2018)。与之类似,前寒武系鲕粒的微生物成因和有机矿化机制在其圈层形成过程中的作用也得到了证实(Diaz et al.,2017;Batchelor,2018;Diaz and Eberli,2019;梅冥相,2021;李飞等,2022)。此外,针对微生物岩的成储研究也取得了丰富的成果。目前认为微生物岩储集空间的发育受控于沉积环境、原始组构、岩溶改造、白云石化等(沈安江等,2022)。然而,这是对所有类型碳酸盐岩都适用的共性机制,关于更能体现微生物岩成岩成储特殊性的讨论,尤其是对典型实例的剖析还有待深入。
微生物不仅能够参与很多碳酸盐岩的建造而形成微生物岩,还可能对微生物岩形成后的成岩改造与孔洞发育产生一定影响,直接关系到微生物岩的成储。基于此,本文对鄂尔多斯盆地南缘中元古界蓟县系冯家湾组叠层石和鲕粒岩中与富有机质组构高度相关的白云石及其溶蚀现象展开讨论,解析微生物因素与成岩、成储的关系,以期为深入认识前寒武系部分白云岩成因和相关的微生物岩孔洞发育机制提供参考。
1. 地质背景
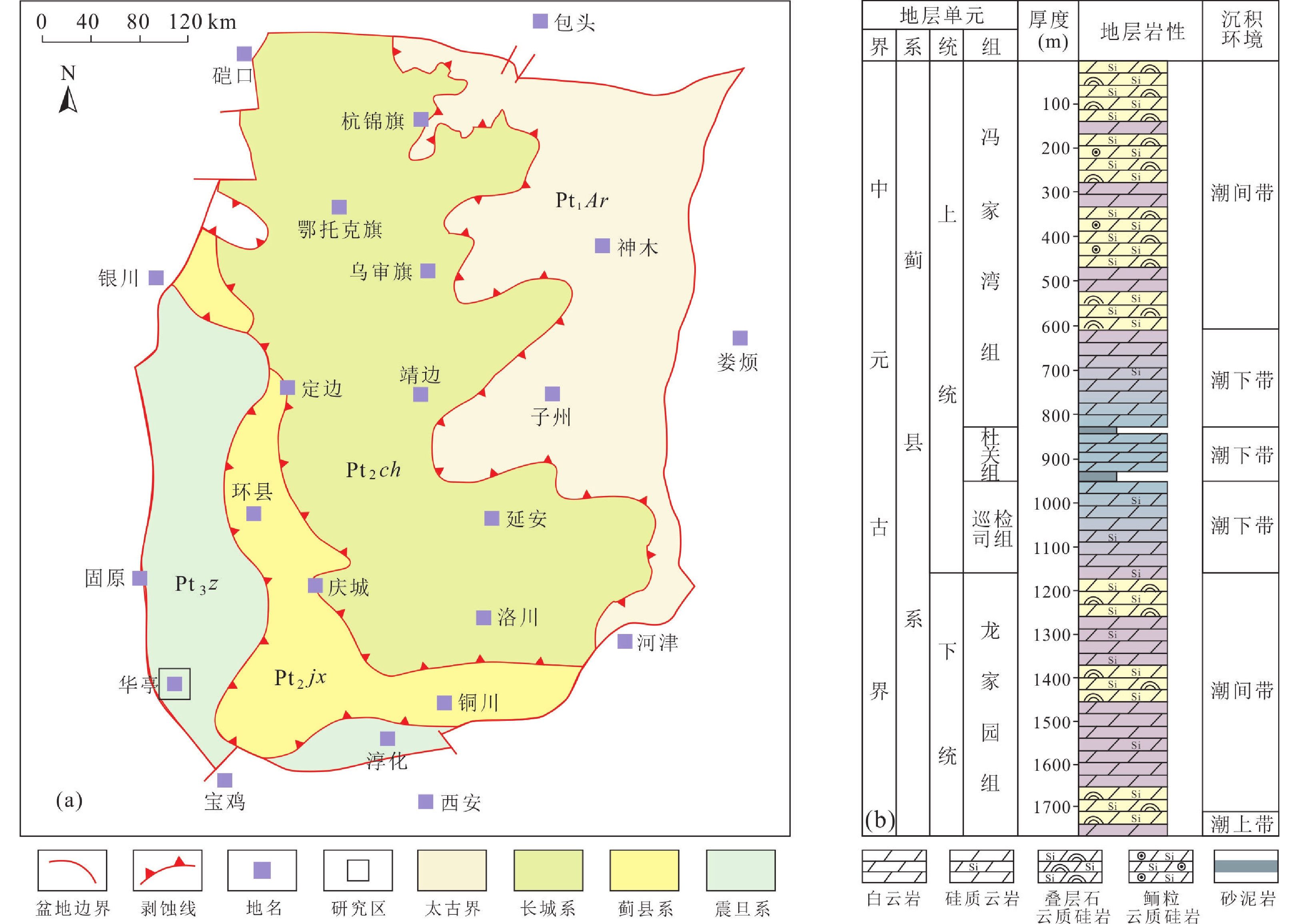
鄂尔多斯盆地是一个发育在太古宙−古元古代变质结晶基底上的多旋回叠合盆地。中元古界长城系和蓟县系是结晶基底上早期沉积盖层的主体。长城系主要为滨海碎屑岩沉积,上覆蓟县系与之为不整合接触。蓟县纪,鄂尔多斯盆地东北部为隆起古陆,西南缘为沉积坳陷区,发育陆表海碎屑岩−碳酸盐岩沉积,构成以盆地西缘王全口群和盆地南缘洛南群为代表的蓟县系,沉积厚度最大可超过
1700 m,由南向北、由西向东超覆,主要分布在陕甘交界和宁夏北部地区(图1a)。其下限年龄距今约1600 Ma(谭聪等,2019)。![]() 图 1 鄂尔多斯盆地前寒武纪地层出露范围(a)与盆地南缘蓟县系沉积层序(b)Figure 1. Distribution (a) and southern sedimentary succession (b) of the Mesoproterozoic Jixian System in the Ordos Basin
图 1 鄂尔多斯盆地前寒武纪地层出露范围(a)与盆地南缘蓟县系沉积层序(b)Figure 1. Distribution (a) and southern sedimentary succession (b) of the Mesoproterozoic Jixian System in the Ordos Basin在盆地南缘,蓟县系由老到新依次为龙家园组、巡检司组、杜关组及冯家湾组。碳酸盐岩主要发育在龙家园组、巡检司组和冯家湾组,岩性以云质硅岩、硅质云岩和白云岩为主。其中,巨厚的冯家湾组和龙家园组发育有大量叠层石和一些鲕粒岩(图1b),并可见多种与微生物生命活动相关的沉积组构。在甘肃华亭的马峡剖面,冯家湾组出露连续,叠层石累计厚度近500 m,沉积、成岩现象典型,是开展研究的优选对象。冯家湾组沉积期距今约
1440 Ma(李怀坤等,2014),处于叠层石发育的高峰期。2. 岩石学特征
叠层石和鲕粒岩是研究区冯家湾组基质孔隙型储集岩的主要岩石类型。X衍射分析显示,二者成分基本由硅质和白云石构成,方解石的含量微乎其微。前寒武纪频繁而强烈的海底火山和热水喷气活动导致当时的海洋中有丰富的可溶SiO2(李延河等,2010),是前寒武系碳酸盐岩硅质含量显著高于显生宙水平的一个重要原因。在这种古海洋环境的影响下,研究区叠层石和鲕粒岩中都含有大量硅质成分。硅质大规模广泛分布,并非主要以条带、结核或团块的形式产出。鲕粒岩的硅质含量比叠层石的更高,前者接近70%,后者多在50%~60%。硅质叠层石和鲕粒岩的基质部分总体较为致密,因局部发育组构选择性孔洞而成储。
研究区叠层石以波状—微波状、层状为主(图2a-b),主要形成于水体能量较低的潮间带上部,具有清晰的明暗相间的纹层结构,发育由富有机质的暗层和由硅质构成的亮层所组成的交替纹层(图2b-c)。纹层厚度变化范围非常大,如暗纹层为0.05~2 mm不等。暗层周缘常发育一种具独特环带结构的“皮壳—葡萄状”胶结物(图2d),很多研究证实其形成离不开微生物的参与(钱一雄等,2017;林孝先等,2018;任冠雄,2018)。叠层石内部夹杂有规模不等的鲕粒岩(图2e),局部含砂屑、砾屑,颗粒间亮晶胶结,反映沉积环境水体能量较高。鲕粒直径多为300~500 µm,类型多样,以同心—放射状真鲕最为常见(图2f)。其圈层结构与叠层石的纹层结构相似,相当于纹层闭合的包覆体。暗圈层中富含有机质,亮圈层为硅质层。单个圈层厚度多为30~50 µm。
![]() 图 2 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩发育特征与相关组构a. 层状硅质叠层石手标本;b. 单偏光下硅质叠层石中的亮暗纹层;c. 正交偏光下的硅质叠层石;d. 单偏光下叠层石暗纹层周缘的“皮壳—葡萄状”胶结物; e. 正交偏光下的硅质鲕粒岩; f. 单偏光下硅质同心放射状真鲕Figure 2. Characteristics and fabrics of the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
图 2 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩发育特征与相关组构a. 层状硅质叠层石手标本;b. 单偏光下硅质叠层石中的亮暗纹层;c. 正交偏光下的硅质叠层石;d. 单偏光下叠层石暗纹层周缘的“皮壳—葡萄状”胶结物; e. 正交偏光下的硅质鲕粒岩; f. 单偏光下硅质同心放射状真鲕Figure 2. Characteristics and fabrics of the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin叠层石和鲕粒中的暗色富有机质组构为微生物富集的组构,其主要组成为微生物膜,系由微生物通过自身分泌的胞外聚合物粘结而成的一种膜状组织(Riding,2000)。前寒武系碳酸盐岩中的微生物体经历漫长地质时期后被保存下来并被直接观察到的难度非常大,更多被观察到的是诸如微生物膜的褶皱、破裂收缩等微生物成因的沉积构造。
3. 白云石与相关孔洞特征
3.1 暗色组构内的白云石晶体
在高倍显微镜和阴极发光下可以观察到大量自形白云石晶体集中分布于硅质叠层石的暗色纹层和硅质鲕粒的暗色圈层处,亮层中少见,具有鲜明的组构选择性(图3a-b)。扫描电镜下还能观察到富有机质组构中微小的菱面体(图3c)。叠层石的微量元素原位成像分析结果显示,暗纹层中的镁元素相对更为富集,呈现出橘红色高值(图3d)。
![]() 图 3 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的白云石发育与分布特征a. 单偏光下硅质叠层石暗纹层中的自形白云石晶体;b. 阴极发光下沿硅质鲕粒圈层分布的白云石;c. 扫描电镜下富有机质组构中的白云石晶体;d. 硅质叠层石中的Mg元素含量原位成像分析图;e. 大面积拼接成像下白云石晶体周缘的缝隙;f. 扫描电镜下自形白云石晶体周缘与硅质之间的缝隙和铁锰氧化物Figure 3. Characteristics of the dolomite crystals in the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
图 3 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的白云石发育与分布特征a. 单偏光下硅质叠层石暗纹层中的自形白云石晶体;b. 阴极发光下沿硅质鲕粒圈层分布的白云石;c. 扫描电镜下富有机质组构中的白云石晶体;d. 硅质叠层石中的Mg元素含量原位成像分析图;e. 大面积拼接成像下白云石晶体周缘的缝隙;f. 扫描电镜下自形白云石晶体周缘与硅质之间的缝隙和铁锰氧化物Figure 3. Characteristics of the dolomite crystals in the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin此外,白云石晶体的大小也有受控于暗色组构规模的倾向。叠层石的纹层规模总体上要比鲕粒的圈层规模大,大部分暗色纹层相对更宽,在其中更容易观察到个体较大的白云石晶体,最大可达150 µm;相比之下,鲕粒的暗色圈层要细得多,其中发育的大部分白云石也明显偏小,多为5~20 µm。在扫描电镜下可以观察到白云石晶体边缘普遍存在着相对规则的缝隙,构成了一定的储集空间(图3e-f),晶体周缘还普见铁锰氧化物(图3f)。
3.2 暗色组构内的溶蚀孔洞
研究区叠层石和鲕粒岩的主力储集空间为基质孔隙,突出表现为分布在暗色富有机质纹(圈)层中的规模不等的孔洞,在低倍显微镜下观察时就像其中的有机质腐烂消失了一样。一些是沿鲕粒边缘和内部暗色圈层分布的微孔,其规模基本受控于鲕粒圈层的厚度,多数为几十微米(图4a-b)。还有一些是分布在叠层石暗色纹层中的微孔与溶孔。由于叠层石的暗色纹层相对较宽,导致其中的孔洞大小悬殊,最大一般不超过2 mm(图4c)。
![]() 图 4 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的溶蚀孔洞发育与分布特征a. 单偏光下鲕粒边缘的微孔,蓝色铸体薄片;b. 单偏光下沿鲕粒圈层分布的微孔,蓝色铸体薄片;c. 单偏光下叠层石暗色纹层中的微孔与溶孔,蓝色铸体薄片;d. 单偏光下图c黄色框区域中的白云石铸模孔、溶蚀扩大孔与部分残余晶体,蓝色铸体薄片;e. 大面积拼接成像下的白云石相关溶蚀孔洞;f. 扫描电镜下发生部分溶蚀的白云石晶体Figure 4. Characteristics of the pores and vugs in the stromatolite and oolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
图 4 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的溶蚀孔洞发育与分布特征a. 单偏光下鲕粒边缘的微孔,蓝色铸体薄片;b. 单偏光下沿鲕粒圈层分布的微孔,蓝色铸体薄片;c. 单偏光下叠层石暗色纹层中的微孔与溶孔,蓝色铸体薄片;d. 单偏光下图c黄色框区域中的白云石铸模孔、溶蚀扩大孔与部分残余晶体,蓝色铸体薄片;e. 大面积拼接成像下的白云石相关溶蚀孔洞;f. 扫描电镜下发生部分溶蚀的白云石晶体Figure 4. Characteristics of the pores and vugs in the stromatolite and oolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin对鲕粒暗色圈层和叠层石暗色纹层内的微孔进行高倍观察时发现,这些微孔主要表现为硅质背景下的菱形白云石晶体铸模孔和在其基础之上形成的溶蚀扩大孔(图4d-e)。铸模孔的大小与暗色组构的规模大致成正比,叠层石中的铸模孔总体大于鲕粒中的铸模孔。局部可见未溶蚀和部分溶蚀的白云石晶体呈漂浮状位于铸模孔或溶孔的中央(图4d),还可见发生部分溶蚀的晶体内部的一些组构选择性溶蚀特征(图4f)。部分相邻的铸模孔相互连通,形成较大的形状欠规则的溶孔。这些铸模孔及其溶蚀扩大孔的组构选择性明确,完全游离于暗色组构之外的亮层中的类似孔隙较少见,这一点在纹层规模更大的叠层石中表现得更为清晰。
4. 白云石的形成与溶蚀
4.1 白云石的发育
暗色富有机质组构内白云石的发育对成储有明显的建设性作用。无论是其形成过程中在晶体周缘出现的缝隙还是晶体溶蚀后形成的孔洞,均对研究区硅质叠层石和鲕粒岩基质部分的储集物性起到了显著的改善作用,是储集空间发育的基础和关键。由于白云石无论是分布特征还是晶体大小均受控于暗色组构,加之周围致密的硅质可在一定程度上阻碍外源性成岩流体的干扰,因此这期白云石的发育可能受到了暗色组构内部微生物因素的影响。
越来越多的研究表明,微生物不能直接诱导沉淀具有菱面体晶形的严格意义上的白云石,但可以通过对微环境中Ca2+、Mg2+的富集和碱度的升高促进高镁方解石、无序或部分有序的富钙白云石等白云石前驱矿物的沉淀(张亦凡等,2015;许杨阳等,2018;赵东方等,2022),并为其在成岩环境中进一步发生有序化调整创造有利条件,从而形成有序度较高的真正意义上的白云石。
首先,微生物的胞外聚合物(extracellular polymeric substances, EPS)含有丰富的具较大比表面积、带负电荷的官能团,可高效吸附Ca2+、Mg2+等金属阳离子并在后期降解时将其释放从而提高局部Ca2+、Mg2+的浓度,为富镁碳酸盐矿物的成核和生长提供必需的物质和场所(图5a;王璇等,2024;钟怡江等,2022)。其次,微生物的生命活动能影响细胞邻近微区的水化学条件,通过升高碱度的方式增加碳酸盐的饱和度,促其发生沉淀。例如,蓝细菌的光合作用能通过二氧化碳浓缩机制吸收CO2和/或HCO3-,增加细胞周围的pH值(图5b;Frankel and Bazylinski,2003;Riding,2006;Dupraz et al.,2009)。即便是在蓝细菌死亡后的降解阶段,仍会发生一些升高碱度的反应(Kenward et al.,2013;Petrash et al.,2017)。在研究区,通过扫描电镜可以观察到叠层石富有机质组构内的一些瘤状矿化产物(图6a)。
![]() a. EPS吸附Ca2+、Mg2+等金属阳离子并提供矿物成核点位示意图;b. 二氧化碳浓缩机制提高环境碱度并促进碳酸钙沉淀模式图;c. 特定官能团破坏Mg2+水合作用能量壁垒模式图;d. 叶绿素a结构示意图Figure 5. Diagram of the mechanism related to microbial-influenced dolomite formation (modified after Riding, 2006; Zhang, 2016; Dong, 2020; Li et al., 2022; Zhao et al., 2022)
a. EPS吸附Ca2+、Mg2+等金属阳离子并提供矿物成核点位示意图;b. 二氧化碳浓缩机制提高环境碱度并促进碳酸钙沉淀模式图;c. 特定官能团破坏Mg2+水合作用能量壁垒模式图;d. 叶绿素a结构示意图Figure 5. Diagram of the mechanism related to microbial-influenced dolomite formation (modified after Riding, 2006; Zhang, 2016; Dong, 2020; Li et al., 2022; Zhao et al., 2022)![]() 图 6 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组硅质叠层石基质孔隙演化综合模式图a. 扫描电镜下叠层石暗纹层内的瘤状矿化产物;b. 扫描电镜下叠层石暗纹层内的白云石晶体及其周缘的铁锰氧化物;c. 扫描电镜下叠层石暗纹层内的白云石晶体铸模孔及其中的硅质沉淀;d. 与富有机质组构相关的白云石形成与溶蚀过程模式图;e. 图d黄色框中的孔隙演化过程模式图Figure 6. Diagrams of the matrix pore space evolution in the siliceous stromatolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
图 6 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组硅质叠层石基质孔隙演化综合模式图a. 扫描电镜下叠层石暗纹层内的瘤状矿化产物;b. 扫描电镜下叠层石暗纹层内的白云石晶体及其周缘的铁锰氧化物;c. 扫描电镜下叠层石暗纹层内的白云石晶体铸模孔及其中的硅质沉淀;d. 与富有机质组构相关的白云石形成与溶蚀过程模式图;e. 图d黄色框中的孔隙演化过程模式图Figure 6. Diagrams of the matrix pore space evolution in the siliceous stromatolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin镁对白云石的发育尤为关键。除了EPS对Mg2+的吸附效应外,EPS中的羧酸盐有机物质与细胞壁中的羧基还具有使镁脱水的功能,可以通过取代镁水合离子中一个或多个水分子的方式降低Mg2+水合作用的能量壁垒(图5c),从而提高参与反应的Mg2+的数量(Petrash et al.,2017;肖恩照等,2017;钟怡江等,2022)。此外,蓝细菌的叶绿素a中也含有镁(图5d;董陈松,2020)。这些因素都会造成富有机质组构内Mg2+含量的增高与活性的增强,既有利于提高有机矿化产物中镁的比例,使其后期更容易向白云石转化,又有利于富镁成岩流体的形成,促进云化的发生。
基于上述因素,研究区虽然存在有利于硅质形成的地质背景,但富有机质暗色组构内部为偏碱性且富镁的微环境,为白云石的发育创造了有利条件。白云石晶体周缘与硅质之间的缝隙、伴随白云石出现的铁锰氧化物以及叠层石暗色纹层中的Mg元素高值都是这一有利成岩环境的反映(图3d,3f,图6b),间接体现了微生物因素对白云石发育的促进作用,由此导致暗色组构中大量白云石的形成。在微生物繁盛并参与造岩的前寒武纪,类似成因的白云岩的规模可能不容小觑。
4.2 溶蚀孔洞的形成
暗色组构中的部分白云石发生了溶蚀,形成晶体铸模孔。在铸模孔发育处,先前白云石晶体周缘的铁锰氧化物全部消失,还可见一些硅质在铸模孔中发生沉淀(图6c),说明富有机质组构内部的成岩微环境已由之前的偏碱性转变为偏酸性。由于致密硅质对外源流体的阻碍,溶蚀作用的发生可能更多地受到埋藏阶段微生物岩中一些能够产生酸性物质的生物化学反应的影响。这些反应能降低富有机质组构内微环境中的pH值(Bengtsson et al.,2008;游杰等,2020),导致白云石发生溶解,主要得益于岩石中较高的微生物有机质丰度。
早成岩阶段的有机质降解是产酸的一个重要途径。例如,蓝细菌在发酵过程中,其中的蛋白质、碳水化合物和脂类会被厌氧微生物最终分解为乙酸、丙酸、丁酸等有机酸,并伴生CO2,溶于水后形成碳酸(李子阳,2020):
$$ \mathrm{有机物} \to {\mathrm{RCOOH+H}}_{ \mathrm{2}} \mathrm{O+CO}_{ \mathrm{2}} \mathrm{,H}_{ \mathrm{2}} \mathrm{O+CO}_{ \mathrm{2}} \to \mathrm{H}_{ \mathrm{2}} \mathrm{CO}_{ \mathrm{3}} \mathrm{。} $$ 此外,微生物有机质在热解生烃过程中也会释放出有机酸和CO2。这些酸性物质与白云石反应后导致其溶解成孔:
$$ {\mathrm{CaMg(CO}_{ \mathrm{3}} \mathrm{)}_{ \mathrm{2}} \mathrm{+RCOOH} \to {\mathrm{RCOO}}^{ \mathrm{-}} \mathrm{+Ca}^{ \mathrm{2+}} \mathrm{+Mg}^{ \mathrm{2+}} \mathrm{+H}_{ \mathrm{2}} \mathrm{O+CO}_{ \mathrm{2}} \mathrm{,}} $$ $$ \mathrm{CaMg(CO}_{ \mathrm{3}} \mathrm{)}_{ \mathrm{2}} \mathrm{+H}_{ \mathrm{2}} \mathrm{CO}_{ \mathrm{3}} \to \mathrm{Ca}^{ \mathrm{2+}} \mathrm{+Mg}^{ \mathrm{2+}} \mathrm{+HCO}_{ \mathrm{3}}^{ \mathrm{-}} \mathrm{。} $$ 现代沉积和模拟实验均表明,微生物有机质的早期低温降解和晚期热解生烃有利于碳酸盐岩次生孔隙的形成和在埋藏环境中的保存(沈安江等,2022)。研究对象中白云石晶体与周缘硅质之间的缝隙则为白云石后期的溶蚀提供了有利的流体通道。
由于暗色富有机质组构内部的微孔隙中也均为硅质,因此在高倍单偏光下,白云石溶蚀后表现为硅质中的铸模孔。在白云石分布较密集且后期溶蚀作用较强烈的区域,铸模孔之间连通后形成溶孔,部分溶孔可进一步贯通形成小洞,最终导致在暗色组构内发育规模不等的溶蚀孔洞,使微生物岩的储集性在局部得到明显改善。因此,暗色组构中白云石的溶蚀是孔洞发育的关键,白云石铸模孔成为冯家湾组硅质叠层石和鲕粒岩基质孔隙的基本构成单元(图6d-e)。由于具有鲜明的组构选择性,在外观上容易造成一种有机质腐烂消失后直接造孔的假象,这是在以往研究中对类似现象形成的习惯性的认识。
然而,受到蓝细菌细胞壁中较难降解并影响细胞中有机质分离释放的生物聚合物等因素的影响(李子阳,2020),上述产酸过程不一定会在所有暗色组构中都顺利发生。微生物有机质中可能只有一部分发生了降解产酸和/或热解产酸,还有一部分始终未能产酸。因此,同一组构中不同微区域的溶蚀强度存在明显差异,不是所有的白云石都能溶蚀成孔,还有很多被完好地保存了下来,由此导致富有机质组构内的孔洞发育具有明显的非均质性。
孔洞形成后虽经历了漫长的地质时期,但大部分仍以未充填和少量被硅质半充填的状态保持至今。一方面与硅质对后期可能充填孔洞的外源成岩流体的阻碍有关。另一方面,由于白云石从流体中直接沉淀下来的难度较大,因此其发生溶蚀后不易在异地再次沉淀,加之冯家湾组硅质叠层石和鲕粒岩中方解石的含量极低,从而造成这些孔洞形成后不易被碳酸盐矿物充填破坏。
5. 结论
(1)鄂尔多斯盆地南缘蓟县系冯家湾组硅质叠层石和鲕粒岩暗色富有机质组构中孔洞的发育是叠层石和鲕粒岩基质部分成储的关键。其并非由有机质腐烂直接释放空间所导致,而是由组构内大量白云石晶体发生溶蚀形成的。
(2)富有机质组构内部Ca2+、Mg2+的富集与碱度的升高有利于白云石的形成,而随后产生的酸性物质又导致这些白云石发生溶解。由于成岩微环境相对封闭,白云石的发育和溶蚀更多地与组构内部微生物因素的影响有关。
(3)受微生物因素影响的白云石的形成与溶蚀,对于主要由微生物岩构成且大多经历了强烈破坏性成岩改造的前寒武系古老碳酸盐岩而言,不但是其中部分白云岩发育的一个重要机制,更是一个值得关注的成储机制。
致谢:两位审稿专家和编辑部老师对本文的内容和写作提出了非常宝贵的意见与建议, 在此表示衷心感谢!
-
![]()
图 1 鄂尔多斯盆地前寒武纪地层出露范围(a)与盆地南缘蓟县系沉积层序(b)
Figure 1. Distribution (a) and southern sedimentary succession (b) of the Mesoproterozoic Jixian System in the Ordos Basin
![]()
图 2 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩发育特征与相关组构
a. 层状硅质叠层石手标本;b. 单偏光下硅质叠层石中的亮暗纹层;c. 正交偏光下的硅质叠层石;d. 单偏光下叠层石暗纹层周缘的“皮壳—葡萄状”胶结物; e. 正交偏光下的硅质鲕粒岩; f. 单偏光下硅质同心放射状真鲕
Figure 2. Characteristics and fabrics of the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
![]()
图 3 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的白云石发育与分布特征
a. 单偏光下硅质叠层石暗纹层中的自形白云石晶体;b. 阴极发光下沿硅质鲕粒圈层分布的白云石;c. 扫描电镜下富有机质组构中的白云石晶体;d. 硅质叠层石中的Mg元素含量原位成像分析图;e. 大面积拼接成像下白云石晶体周缘的缝隙;f. 扫描电镜下自形白云石晶体周缘与硅质之间的缝隙和铁锰氧化物
Figure 3. Characteristics of the dolomite crystals in the stromatolite and oolite from the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
![]()
图 4 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组叠层石和鲕粒岩中的溶蚀孔洞发育与分布特征
a. 单偏光下鲕粒边缘的微孔,蓝色铸体薄片;b. 单偏光下沿鲕粒圈层分布的微孔,蓝色铸体薄片;c. 单偏光下叠层石暗色纹层中的微孔与溶孔,蓝色铸体薄片;d. 单偏光下图c黄色框区域中的白云石铸模孔、溶蚀扩大孔与部分残余晶体,蓝色铸体薄片;e. 大面积拼接成像下的白云石相关溶蚀孔洞;f. 扫描电镜下发生部分溶蚀的白云石晶体
Figure 4. Characteristics of the pores and vugs in the stromatolite and oolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
![]()
图 5 受微生物因素影响的白云石成因综合模式图(据Riding,2006;张喜洋,2016;董陈松,2020;李飞等,2022;赵东方等,2022修编)
a. EPS吸附Ca2+、Mg2+等金属阳离子并提供矿物成核点位示意图;b. 二氧化碳浓缩机制提高环境碱度并促进碳酸钙沉淀模式图;c. 特定官能团破坏Mg2+水合作用能量壁垒模式图;d. 叶绿素a结构示意图
Figure 5. Diagram of the mechanism related to microbial-influenced dolomite formation (modified after Riding, 2006; Zhang, 2016; Dong, 2020; Li et al., 2022; Zhao et al., 2022)
![]()
图 6 鄂尔多斯盆地南缘马峡剖面蓟县系冯家湾组硅质叠层石基质孔隙演化综合模式图
a. 扫描电镜下叠层石暗纹层内的瘤状矿化产物;b. 扫描电镜下叠层石暗纹层内的白云石晶体及其周缘的铁锰氧化物;c. 扫描电镜下叠层石暗纹层内的白云石晶体铸模孔及其中的硅质沉淀;d. 与富有机质组构相关的白云石形成与溶蚀过程模式图;e. 图d黄色框中的孔隙演化过程模式图
Figure 6. Diagrams of the matrix pore space evolution in the siliceous stromatolite of the Fengjiawan Formation of Jixian System at Maxia section in southern Ordos Basin
-
[1] Batchelor M T,Burne R V,Henry B I,et al.,2018. A biofilm and organomineralisation model for the growth and limiting size of ooids[J]. Scientific Reports,8(1):559. DOI: 10.1038/s41598-017-18908-4
[2] Bengtsson S,Hallquist J,Werker A,et al.,2008. Acidogenic fermentation of industrial wastewaters:Effects of chemostat retention time and pH on volatile fatty acids production[J]. Biochemical Engineering Journal,40(3):492 − 499. DOI: 10.1016/j.bej.2008.02.004
[3] Bosence D,Gibbons K,Le Heron D P,et al.,2015. Microbial carbonates in space and time:introduction[J]. Geological Society,London,Special Publications,418(1):1 − 15. DOI: 10.1144/SP418.14
[4] Diaz M R,Eberli G P,2019. Decoding the mechanism of formation in marine ooids:A review[J]. Earth-Science Reviews,190:536 − 556. DOI: 10.1016/j.earscirev.2018.12.016
[5] Diaz M R,Eberli G P,Blackwelder P,et al.,2017. Microbially mediated organomineralization in the formation of ooids[J]. Geology,45(9):771 − 774. DOI: 10.1130/G39159.1
[6] 董陈松,2020. 蓝细菌中光依赖性原叶绿素酸酯氧化还原酶的结构和分子机理[D]. 合肥:安徽大学. Dong C S,2020. Structural and functional studies of cyanobacteria light-dependent protochlorophyllide oxidoreductase[D]. Hefei:Anhui University (in Chinese with English abstract).
[7] Dupraz C,Reid R P,Braissant O,et al.,2009. Processes of carbonate precipitation in modern microbial mats[J]. Earth Science Reviews,96:141 − 162. DOI: 10.1016/j.earscirev.2008.10.005
[8] Frankel R B,Bazylinski D A,2003. Biologically induced mineralization by bacteria[J]. Reviews in Mineralogy & Geochemistry,54(1):95 − 114.
[9] 胡安平,沈安江,郑剑锋,等,2021. 微生物碳酸盐岩分类、沉积环境与沉积模式[J]. 海相油气地质,26(1):1 − 15. Hu A P,Shen A J,Zheng J F,et al.,2021. The classification,facies and sedimentary models of microbialites[J]. Marine Origin Petroleum Geology,26(1):1 − 15 (in Chinese with English abstract).
[10] Johnston D T,Wolfe-Simon F,Pearson A,2009. Anoxygenic photosynthesis modulated Proterozoic oxygen and sustained Earth's middle age[J]. Proceedings of the National Academy of Sciences of the United States of America,106(40):16925 − 16929.
[11] Kenward P A,Fowle D A,Goldstein R H,et al.,2013. Ordered low-temperature dolomite mediated by carboxyl-group density of microbial cell walls[J]. AAPG Bulletin,97(11):2113 − 2125. DOI: 10.1306/05171312168
[12] 李飞,易楚恒,李红,等,2022. 微生物成因鲕粒研究进展[J]. 沉积学报,40(2):319 − 334. Li F,Yi C H,Li H,et al.,2022. Recent advances in ooid microbial origin:A review[J]. Acta Sedimentologica Sinica,40(2):319 − 334 (in Chinese with English abstract).
[13] 李怀坤,苏文博,周红英,等,2014. 中−新元古界标准剖面蓟县系首获高精度年龄制约——蓟县剖面雾迷山组和铁岭组斑脱岩锆石SHRIMP U-Pb同位素定年研究[J]. 岩石学报,30(10):2999 − 3012. Li H K,Su W B,Zhou H Y,et al.,2014. The first precise age constraints on the Jixian System of the Meso-to Neoproterozoic Standard Section of China:SHRIMP zircon U-Pb dating of bentonites from the Wumishan and Tieling formations in the Jixian Section,North China Craton[J]. Acta Petrologica Sinica,30(10):2999 − 3012 (in Chinese with English abstract).
[14] 李延河,侯可军,万德芳,等,2010. 前寒武纪条带状硅铁建造的形成机制与地球早期的大气和海洋[J]. 地质学报,84(9):1359 − 1373. Li Y H,Hou K J,Wan D F,et al.,2010. Formation mechanism of Precambrian banded iron formation and atmosphere and ocean during early stage of the earth[J]. Acta Geologica Sinica,84(9):1359 − 1373 (in Chinese with English abstract).
[15] 李子阳,2020. 蓝藻厌氧发酵产酸及发酵液作为反硝化外加碳源研究[D]. 无锡:江南大学. Li Z Y,2020. Acid production from cyanobacteria by anaerobic fermentation and its application as additional carbon source for denitrification[D]. Wuxi:Jiangnan University (in Chinese with English abstract).
[16] 林孝先,彭军,侯中健,等,2018. 四川汉源−峨边地区上震旦统灯影组藻白云岩特征及成因研究[J]. 沉积学报,36(1):57 − 71. Lin X X,Peng J,Hou Z J,et al.,2018. Study on characteristics and geneses of algal dolostone of the Upper Sinian Dengying Formation in the Hanyuan-Ebian area of Sichuan Province,China[J]. Acta Sedimentologica Sinica,36(1):57 − 71 (in Chinese with English abstract).
[17] 马赛,2020. 鄂尔多斯盆地西南缘构造演化与中上元古界成藏条件[D]. 西安:西北大学. Ma S,2020. Tectonic evolution and Middle-Upper Proterozoic petroleum geological setting in south-western Ordos Basin[D]. Xi'an:Northwest University (in Chinese with English abstract).
[18] 梅冥相,2021. 光合作用生物膜诱发的放射鲕粒:以江苏徐州贾旺剖面寒武系苗岭统张夏组为例[J]. 古地理学报,23(3):461 − 488. Mei M X,2021. Radial ooids induced by photosynthetic biofilms:An example from the Cambrian Miaolingian Zhangxia Formation at Jiawang section in Xuzhou city of Jiangsu Province,North-China Platform[J]. Journal of Palaeogeography,23(3):461 − 488 (in Chinese with English abstract).
[19] Pace A,Bourillot R,Bouton A,et al.,2018. Formation of stromatolite lamina at the interface of oxygenic-anoxygenic photosynthesis[J]. Geobiology,16(4):378 − 398. DOI: 10.1111/gbi.12281
[20] Peters S E,Husson J M,Wilcots J,2017. The rise and fall of stromatolites in shallow marine environments[J]. Geology,45:487 − 490.
[21] Petrash D A,Bialik O M,Bontognali T R R,et al.,2017. Microbially catalyzed dolomite formation:From near-surface to burial[J]. Earth-Science Reviews,171:558 − 582. DOI: 10.1016/j.earscirev.2017.06.015
[22] 钱一雄,冯菊芳,何治亮,等,2017. 从岩石学及微区同位素探讨四川盆地灯影组皮壳−葡萄状白云石成因[J]. 石油与天然气地质,38(4):665 − 676. Qian Y X,Feng J F,He Z L,et al.,2017. Applications of petrography and isotope analysis of micro-drill samples to the study of genesis of grape-like dolomite of the Dengying Formation in the Sichuan Basin[J]. Oil & Gas Geology,38(4):665 − 676 (in Chinese with English abstract).
[23] Reid R P,Visscher P T,Decho A W,et al.,2000. The role of microbes in accretion,lamination and early lithification of modern marine stromatolites[J]. Nature,406:989 − 992. DOI: 10.1038/35023158
[24] 任冠雄,2018. 四川盆地震旦系灯影组葡萄状构造精细研究[D]. 成都:西南石油大学. Ren G X,2018. Study on the botryoidal structure of the Sinian Dengying Formation in Sichuan Basin[D]. Chengdu:Southwest Petroleum University (in Chinese with English abstract).
[25] Riding R,1991. Calcareous algae and stromatolites[M]. Berlin:Springer-Verlag:21 − 51.
[26] Riding R,2000. Microbial carbonates:The geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology,47(1):179 − 214.
[27] Riding R,2006. Cyanobacterial calcification,carbon dioxide concentrating mechanisms,and Proterozoic-Cambrian changes in atmospheric composition[J]. Geobiology,4(4):299 − 316. DOI: 10.1111/j.1472-4669.2006.00087.x
[28] Riding R,2008. Abiogenic,microbial and hybrid authigenic carbonate crusts:Components of Precambrian stromatolites[J]. Geologia Croatica,61(2 − 3):73 − 103.
[29] 沈安江,胡安平,张杰,等,2022. 微生物碳酸盐岩“三因素”控储地质认识和分布规律[J]. 石油与天然气地质,43(3):582 − 596. Shen A J,Hu A P,Zhang J,et al.,2022. "Three-factor" driven microbial carbonate reservoirs and their distribution[J]. Oil & Gas Geology,43(3):582 − 596 (in Chinese with English abstract).
[30] 谭聪,卢远征,宋昊南,等,2019. 华北克拉通西南缘高山河组凝灰岩锆石U-Pb年龄及其地质意义[J]. 地质学报,93(5):1113 − 1124. Tan C,Lu Y Z,Song H N,et al.,2019. Zircon U-Pb dating of the Gaoshanhe Formation tuff in the southwestern margin of the North China craton,and its geological significance[J]. Acta Geological Sinica,93(5):1113 − 1124 (in Chinese with English abstract).
[31] 王璇,梅朝佳,李忠日,2024. 寒武系张夏组树形石中的钙化蓝细菌——以山东淄博峨庄镇后紫峪剖面为例[J]. 沉积与特提斯地质,44(2):339 − 350. Wang X,Mei C J,Li Z R,2024. Calcifying cyanobacteria in dendrolites of the Cambrian Zhangxia Formation: A case study of the Houziyu section in Ezhuang Town,Zibo City,Shandong Province[J]. Sedimentary Geology and Tethyan Geology,44(2):339 − 350 (in Chinese with English abstract).
[32] 肖恩照,隋明园,Latif K,等,2017. 微生物白云岩形成机制研究进展与存在问题[J]. 大庆石油地质与开发,36(2):26 − 32. Xiao E Z,Sui M Y,Latif K,et al.,2017. Study advances and existed problem for the forming mechanism of the microbial dolomite[J]. Petroleum Geology & Oilfield Development in Daqing,36(2):26 − 32 (in Chinese with English abstract).
[33] 许杨阳,刘邓,于娜,等,2018. 微生物(有机)白云石成因模式研究进展与思考[J]. 地球科学,43(Suppl1):63 − 70. Xu Y Y,Liu D,Yu N,et al.,2018. Advance and review on microbial/organogenic dolomite model[J]. Earth Science,43(Suppl1):63 − 70 (in Chinese with English abstract).
[34] 游杰,胡广,张玺华,等,2020. 微生物碳酸盐岩同生−早成岩阶段有机质降解示踪:以四川盆地灯影组四段为例[J]. 南京大学学报:自然科学版,56(3):308 − 321. You J,Hu G,Zhang X H,et al.,2020. Geochemical tracing of organic matter degradation in microbial carbonates during syngenetic-early diagenesis:A case study from the Member IV of Dengying Formation,Sichuan Basin[J]. Journal of Nanjing University (Natural Sciences),56(3):308 − 321 (in Chinese with English abstract).
[35] 张喜洋. 2016. 豫西寒武纪微生物碳酸盐岩中蓝细菌的钙化作用[D]. 焦作:河南理工大学. Zhang X Y,2016. The calcification of cyanobacteria from microbial carbonates in the Cambrian,western Henan[D]. Jiaozuo:Henan Polytechnic University (in Chinese with English abstract).
[36] 张亦凡,马怡飞,姚奇志,等,2015. “白云石问题”及其实验研究[J]. 高校地质学报,21(3):395 − 406. Zhang Y F,Ma Y F,Yao Q Z,et al.,2015. "Dolomite problem" and experimental studies of dolomite formation[J]. Geological Journal of China Universities,21(3):395 − 406 (in Chinese with English abstract).
[37] 赵东方,谭秀成,罗冰,等,2022. 微生物诱导白云石沉淀研究进展及面临的挑战[J]. 沉积学报,40(2):335 − 349. Zhao D F,Tan X C,Luo B,et al.,2022. A review of microbial dolomite:Advances and challenges[J]. Acta Sedimentologica Sinica,40(2):335 − 349 (in Chinese with English abstract).
[38] 钟怡江,文华国,陈洪德,等,2022. 胞外聚合物在蓝细菌钙化过程中的作用及其地质意义[J]. 沉积学报,40(1):88 − 105. Zhong Y J,Wen H G,Chen H D,et al.,2022. The role of extracellular polymeric substances in cyanobacterial calcification and its geological significance[J]. Acta Sedimentologica Sinica,40(1):88 − 105 (in Chinese with English abstract).
 下载:
下载:






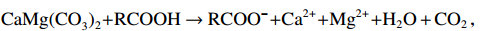

计量
- 文章访问数: 10
- HTML全文浏览量: 0
- PDF下载量: 4
